1. Active in vivo translocation of the Methanosarcina mazei Gö1 Casposon. Nucleic Acids Res. 2023 2. Casposase structure and the mechanistic link between DNA transposition and spacer acquisition by CRISPR-Cas eLife 9:e50004.
07-04-2025
3. Addy
Transposition of Mutator-like transposable elements (MULEs) resembles hAT and Transib elements and V(D)J recombination-2017-NAR
21-04-2025
4. Wijdan
A Naturally Active Spy Transposon Discovered from the Insect Genome of Colletes gigas as a Promising Novel Gene Transfer Tool-2024-ADS
28-04-2025
5. Ahmed
Characterizing Mobile Element Insertions in 5,675 Genomes
6-5-2025
6. XXxing
All-RNA-mediated targeted gene integration in mammalian cells with rationally engineered R2 retrotransposons-cell
19-05-2025
7. XianKL
Cas9-induced targeted integration of large DNA payloads in primary human T cells via homology-mediated end-joining DNA repair-2024-nbe
2-6-2025
8. LiMY
Transposase-assisted target-site integration for efficient plant genome engineering-2024-nature
9-06-2025
9. WanQ
Transposase N-terminal phosphorylation and asymmetric transposon ends inhibit piggyBac transposition in mammalian cells-NAR-2022
16-06-2025
1. Phiri
Genomic Analysis Revealed a Convergent Evolution of LINE-1 in Coat Color_ A Case Study in Water Buffaloes (Bubalus bubalis)-MBE-2021
23-06-2025
2. XinChen
Megabase-scale human genome rearrangement with programmable bridge recombinases-2025-preprint
30-06-2025
3. Addy
Functional characterization of the active Mutator- like transposable element, Muta1 from the mosquito Aedes aegypti-mobile DNA-2017
July-14
4. XianKL
Protein Mutational Effect Predictor (ProMEP) Zero-shot prediction of mutation effects with multimodal deep representation learning guides protein engineering
July-28
5. Ahmed
An Alu insertion map of the Indian population: identification and analysis in 1021 genomes of the IndiGen project. NAR Genom Bioinform. 2022
August-11
6. NSYang
TnMining protocol
1-Sep.
7. XXxing
Structure and biochemistry-guided engineering of an all-RNA system for DNA insertion with R2 retrotransposons
8-Sep.
8. Wijdan
Transposase expression, element abundance, element size, and DNA repair determine the mobility and heritability of PIF-Pong-Harbinger transposable elements-2023-front-cell-and-biology
15-Sep.
9. LiMY
MiCas9 increases large size gene knock-in rates and reduces undesirable on-target and off-target indel edits-2020-nc
10. WanQ
Discovery and structural mechanism of DNA endonucleases guided by RAGATH-18-derived RNAs-2024
10.
A programmable seekRNA guides target selection by IS1111 and IS110 type insertion sequences
Latent activity in TnpB revealed by mutational scanning-2025
PPT in English and Chinese, talk less than 15 minutes in Eng.
Transposases can be classified based on the mechanism of transposition, sequence specificity, and structural features.
1. DDE Transposases (RNase H-like transposases):
These transposases are named after a characteristic triad of acidic residues (Asp-Asp-Glu) in their catalytic domain. This family includes several well-studied transposases.
2. Tyrosine Transposases (Y1):
These typically use a tyrosine residue for transesterification reactions that facilitate transposition. They often exhibit some similarity in mechanism to site-specific recombinases, which also use tyrosine for catalysis.
3. Serine Transposases:
These transposases utilize a serine residue in their catalytic mechanism. Like other serine recombinases, they form a covalent protein-DNA intermediate during transposition.
4. HUH single-stranded DNA transposases (Rolling Circle (RC) Transposases, or Y2):
Also known as IS91-like transposases, these enzymes use a rolling-circle mechanism of transposition. Unlike the cut-and-paste mechanism, RC transposition involves replication of the transposable element at the new site while the original remains in place.
5. Helitron Transposases:
Helitron transposons transpose by a rolling circle mechanism that is distinct from both classic DNA transposons and typical RC transposases. They are often studied in plants, where they play a significant role in genome evolution.
6. Transib Transposases:
This lesser-known class shares some features with both DDE and rolling-circle transposases but is distinguished by unique structural and sequence features. Transib transposases are found in a variety of organisms and are thought to have been involved in the evolution of the RAG recombinase crucial for vertebrate immune system function.
7. Reverse transcriptases/endonucleaseses (RT/En): combination of reverse transcriptase and endonuclease activities Each class of transposases has evolved unique mechanisms and structural adaptations that allow them to facilitate the movement of their associated transposable elements. Their diversity reflects the wide range of roles they play in genomic rearrangement and evolution across different species.
DNA转座子类型
剪切-粘贴型DNA转座子/Cut-and-Paste transposon
CUT-AND-PASTE DNA TRANSPOSONS AS GENETIC TOOLS Cut-and-paste DNA transposons are mobile genetic elements that can move from one genomic location to another through a process known as transposition. They “cut” themselves out of one location in the genome and “paste” themselves into a new location. This capability makes them an invaluable tool for genetic studies and biotechnological applications. Here, we explore the use of cut-and-paste DNA transposons as genetic tools.
Mechanism of Action The classic cut-and-paste DNA transposons move through a “cut-and-paste” mechanism, which involves the following steps: 1. Recognition: The transposase enzyme, encoded by the transposon, binds to specific sequences at the ends of the transposon. 2. Excision: The enzyme cuts the DNA at these points, excising the transposon from its original location. 3. Integration: The transposase then mediates the integration of the transposon at a new site in the genome.
This process can occur without the need for DNA replication, which differs from the replication-based movement seen in some other types of transposable elements.
Applications as Genetic Tools Cut-and-paste DNA transposons are used in a variety of ways in genetic research and biotechnology:
Gene Delivery: Transposons can be used to insert genes into cells, making them useful for gene therapy or creating transgenic organisms.
Mutagenesis: By inserting themselves into or near genes, transposons can disrupt gene function, helping researchers study gene roles and create model organisms with specific mutations.
Genome Manipulation: Transposons can be used to create insertions, deletions, and inversions in the genome, allowing for large-scale modifications and functional genomics studies.
Gene Tagging: Transposons can serve as molecular tags to label genes. This is particularly useful in forward genetics, where a phenotype is traced back to the responsible gene.
Transgenic Tool: They can be employed to stably integrate transgenes into the genome, which is essential in creating transgenic plants and animals for agriculture and research.
Functional Genomics: Transposons can be used for gene knockout libraries, enabling the systematic analysis of gene function.
Controlling Transposition To harness transposons as tools, scientists have developed methods to control their activity: – Binary Systems: Use two components, one to express the transposase and another containing the transposable element, to control spatial and temporal transposition. – Inducible Promoters: These can regulate the expression of transposase, activating transposition only in the presence of specific inducers or under certain conditions.
Limitations and Challenges While cut-and-paste DNA transposons are powerful tools, there are limitations and challenges to their use: – Random Insertion: Transposons can insert into random genomic locations, which might cause unintended mutations or disruptions. – Transposon Silencing: Over time, cells may silence transposons, limiting their utility for long-term expression studies. – Efficacy: The efficiency of transposase enzymes can vary, and not all transposons are active in all cell types or species.
In Silico Analysis and Modeling To understand and predict the behavior of transposons, computational models and simulations can be used alongside experimental data. Programming tools and genetic algorithms can help in: – Analyzing transposon insertion patterns across genomes. – Predicting potential off-target effects and assessing risk factors. – Exploring evolutionary dynamics of transposable elements within populations.
Programming can be leveraged to automate these analyses, utilizing languages such as Python or R, combined with bioinformatics tools and databases.
In conclusion, cut-and-paste DNA transposons serve as versatile genetic tools with a wide range of applications in modern biology. However, their use requires careful management and understanding to fully leveraging their capabilities while minimizing unintended consequences. With ongoing advancements in genetic engineering and control mechanisms, the role of transposons as a genetic tool will likely continue to expand and evolve.
复制-粘贴型DNA转座子/Copy-out-paste-in transposon
译自:Copy-out–Paste-in Transposition of IS911: A Major Transposition Pathway
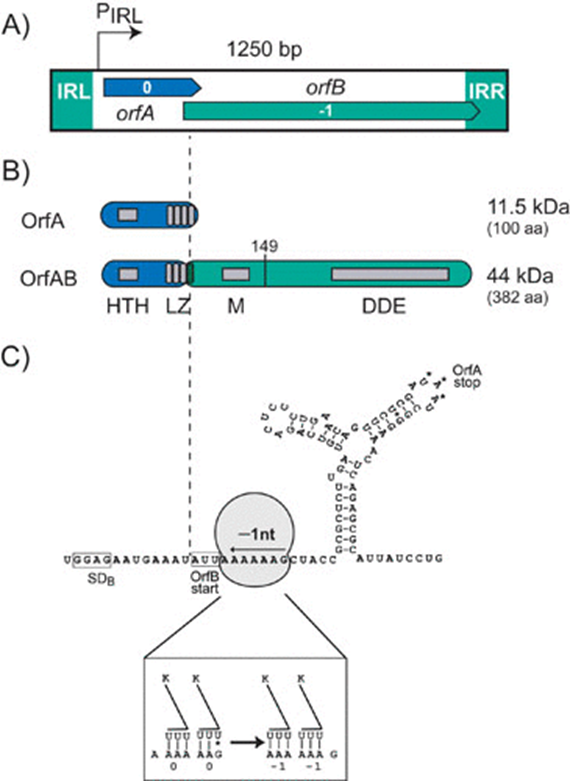
Organization of IS911. (A) Genetic organization. The 1,250-bp IS911 is shown as a box. The boxes at each end represent the left (IRL) and right (IRR) terminal inverted repeats. The two open reading frames, orfA (blue) and orfB (green) are positioned in relative reading phases 0 and −1, respectively, as indicated. The indigenous promoter, pIRL, is shown. The region of overlap between orfA and orfB, which includes the frameshifting signals to produce OrfAB, lies within IS911 coordinates 300 and 400. The precise point at which the frameshift occurs, within the last heptad of the LZ, is indicated by the vertical dotted line. (B) Structure function map of OrfAB and OrfA. HTH, a potential helix-turn-helix motif; LZ, a leucine zipper motif involved in homo- and hetero-multimerization of OrfAB and OrfA. Programmed translational frameshifting that fuses OrfA and OrfB to generate the transposase OrfAB occurs within the fourth heptad. The LZ of OrfA and OrfAB therefore differ in their fourth heptad. A second region, M, necessary for multimerization of OrfAB is shown, as is the catalytic core of the enzyme which carries a third multimerization domain. OrfA translation initiates at an AUG, terminates with UAA whereas OrfAB translation terminates within the right IR. The vertical line to the right of M shows the extent of the truncated transposase, OrfAB[1–149] described in the text. (C) Frameshifting window. The mRNA sequence around the programmed translational frameshifting window is presented. The boxed sequence GGAG is the potential ribosome-binding site located upstream of orfB whose potential translation would be initiated at the boxed AUU codon. A ribosome (not to scale) is shown covering a series of “slippery” codons (AAAAAAG). A downstream secondary structure is also shown with the UAA, OrfA translation termination codon. The ribosome-binding site, slippery codons and secondary structure all contribute to the efficiency of the programmed −1 frameshift. The box at the foot of this figure shows how the anti-codons of two tRNALys are thought to undergo re-pairing with their codons in the AAAAAAG motif.
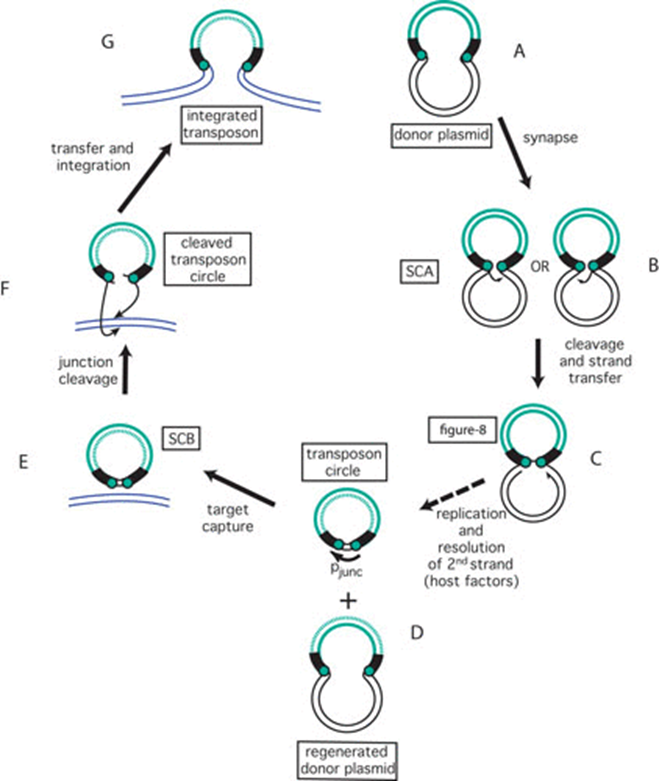
图5. IS911转座循环。 转座子以绿色显示,侧翼供体DNA为黑色,靶DNA为蓝色。转座子末端显示为绿色实心圆。图4中显示的小箭头因简略而省略。(A)携带插入序列(IS)的供体质粒。(B)形成首个突触复合物SCA,切割左侧或右侧的倒置重复序列(IR),并攻击另一端。(C)供体质粒形成单链桥以创建“8”字形分子。(D)IS特异性复制的产物:双链圆形IS转座中间体和再生的转座子供体质粒。复制的链以绿色虚线显示。(E)形成第二个突触复合物SCB并接合靶DNA(蓝色)。(F)切割IS圈并整合。(G)新整合的IS。(The IS911 transposition cycle. The transposon is shown in green, the flanking donor DNA in black and the target DNA in blue. Transposon ends are shown as green filled circles. The small arrows shown in Figure 4 have been omitted for brevity. (A) Donor plasmid carrying the insertion sequence (IS). (B) Formation of the first synaptic complex SCA and cleavage of the left or right inverted repeat (IR) and attack of the other end. (C) Formation of a single-strand bridge to create a figure-eight molecule if the donor is a plasmid as shown here. (D) The products of IS-specific replication: the double strand circular IS transposition intermediate and the regenerated transposon donor plasmid. The replicated strand is shown as a green dotted line. (E) Formation of the second synaptic complex SCB and engagement of the target DNA (blue). (F) Cleavage of the IS circle and integration. (G) The newly integrated IS.)
Transposable elements, contributors in the evolution of organisms (from an arms race to a source of raw materials), Heliyon. 2021. PMID: 33532648.
Specificities and Dynamics of Transposable Elements in Land Plants. Biology (Basel). 2022. PMID: 35453688
A Field Guide to Eukaryotic Transposable Elements. Annu Rev Genet. 2020. PMID: 32955944. A survey of transposable element classification systems–a call for a fundamental update to meet the challenge of their diversity and complexity. Mol Phylogenet Evol. 2015. PMID: 25797922. All Quiet on the TE Front? The Role of Chromatin in Transposable Element Silencing. Cells. 2022. PMID: 36010577. An epigenetic toolkit allows for diverse genome architectures in eukaryotes. Curr Opin Genet Dev. 2015. PMID: 26649755. Bridging multiple dimensions: roles of transposable elements in higher-order genome regulation. Curr Opin Genet Dev. 2023. PMID: 37028152. Coevolution between transposable elements and recombination. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 29109221. Constitutive Heterochromatin in Eukaryotic Genomes: A Mine of Transposable Elements. Cells. 2022. PMID: 35269383. Dark genome, bright ideas: Recent approaches to harness transposable elements in immunotherapies. Cancer Cell. 2022. PMID: 35907399. Distinguishing friends, foes, and freeloaders in giant genomes. Curr Opin Genet Dev. 2018. PMID: 29544119. Diversity and evolution of transposable elements in Arabidopsis. Chromosome Res. 2014. PMID: 4801342. Double-edged sword: The evolutionary consequences of the epigenetic silencing of transposable elements. PLoS Genet. 2020. PMID: 32673310. Epigenetic regulation of intragenic transposable elements: a two-edged sword. J Biochem. 2018. PMID: 30010918. Evolution of transposable elements and evolution of eukaryote genomes mediated by transposable elements. Genes Genet Syst. 2019. PMID: 31996493. Evolutionary impact of transposable elements on genomic diversity and lineage-specific innovation in vertebrates. Chromosome Res. 2015. PMID: 26395902. Giant Transposons in Eukaryotes: Is Bigger Better? Genome Biol Evol. 2019. PMID: 30796812. Horizontal acquisition of transposable elements and viral sequences: patterns and consequences. Curr Opin Genet Dev. 2018. PMID: 29505963. Horizontal transfers of transposable elements in eukaryotes: The flying genes. C R Biol. 2016. PMID: 27234293. Host Gene Regulation by Transposable Elements: The New, the Old and the Ugly. Viruses. 2020. PMID: 32993145. Host-transposon interactions: conflict, cooperation, and cooption. Genes Dev. 2019. PMID: 31481535. How retrotransposons shape genome regulation. Curr Opin Genet Dev. 2016.PMID: 26855260. Impact of transposable elements on the evolution of complex living systems and their epigenetic control. Biosystems. 2021. PMID: 34718084. Integration of TE Induces Cancer Specific Alternative Splicing Events. Int J Mol Sci. 2022. PMID: 36142830. Integration site selection by retroviruses and transposable elements in eukaryotes. Nat Rev Genet. 2017. PMID: 28286338. Investigating the Potential Roles of SINEs in the Human Genome. Annu Rev Genomics Hum Genet. 2021. PMID: 33792357. Jumping the fine LINE between species: horizontal transfer of transposable elements in animals catalyses genome evolution. Bioessays. 2013. PMID: 24003001. Mammalian genome evolution as a result of epigenetic regulation of transposable elements. Biomol Concepts. 2014. PMID: 25372752. Mammalian genome innovation through transposon domestication. Nat Cell Biol. 2022. PMID: 36008480. Mammalian transposable elements and their impacts on genome evolution. Chromosome Res. 2018. PMID: 29392473. Massive contribution of transposable elements to mammalian regulatory sequences. Semin Cell Dev Biol. 2016. PMID: 27174439. Methodologies for the De novo Discovery of Transposable Element Families. Genes (Basel). 2022. PMID: 35456515. Mobility connects: transposable elements wire new transcriptional networks by transferring transcription factor binding motifs. Biochem Soc Trans. 2020. PMID: 32573687. More than causing (epi)genomic instability: emerging physiological implications of transposable element modulation. J Biomed Sci. 2021. PMID: 34364371. New Insights on the Evolution of Genome Content: Population Dynamics of Transposable Elements in Flies and Humans. Methods Mol Biol. 2019. PMID: 31278675. On the Importance to Acknowledge Transposable Elements in Epigenomic Analyses. Genes (Basel). 2019. PMID: 30935103 Recent Bioinformatic Progress to Identify Epigenetic Changes Associated to Transposable Elements. Front Genet. 2022. PMID: 35646069. Regulatory activities of transposable elements: from conflicts to benefits. Nat Rev Genet. 2017. PMID: 27867194 Repetitive elements in aging and neurodegeneration. Trends Genet. 2023. PMID: 36935218. Retrotransposons as a Source of DNA Damage in Neurodegeneration. Front Aging Neurosci. 2022. PMID: 35058771. Rewiring of chromatin state and gene expression by transposable elements. Dev Growth Differ. 2021. PMID: 34050925. Role of Transposable Elements in Gene Regulation in the Human Genome. Life (Basel). 2021. PMID: 33557056. Role of Transposable Elements in Genome Stability: Implications for Health and Disease. Int J Mol Sci. 2022. PMID: 35887150. Roles of Transposable Elements in the Different Layers of Gene Expression Regulation. Int J Mol Sci. 2019. PMID: 31731828. Roles of transposable elements in the regulation of mammalian transcription. Nat Rev Mol Cell Biol. 2022. PMID: 35228718. Silencing of Transposable Elements by piRNAs in Drosophila: An Evolutionary Perspective. Genomics Proteomics Bioinformatics. 2017. PMID:28602845. Structural and sequence diversity of eukaryotic transposable elements. Genes Genet Syst. 2020. PMID: 30416149. Taming the Turmoil Within: New Insights on the Containment of Transposable Elements. Trends Genet. 2020. PMID: 32473745. Taming transposable elements in vertebrates: from epigenetic silencing to domestication. Trends Genet. 2022. PMID: 35307201. Taming, Domestication and Exaptation: Trajectories of Transposable Elements in Genomes. Cells. 2021. PMID: 34944100. TFs for TEs: the transcription factor repertoire of mammalian transposable elements. Genes Dev. 2021. PMID: 33397727. The challenges of predicting transposable element activity in hybrids. Curr Genet. 2021. PMID: 33738571. The evolution and function of transposons in epigenetic regulation in response to the environment. Curr Opin Plant Biol. 2022. PMID: 35961279. The Evolutionary Volte-Face of Transposable Elements: From Harmful Jumping Genes to Major Drivers of Genetic Innovation. Cells. 2021. PMID: 34831175. The impact of transposable elements in genome evolution and genetic instability and their implications in various diseases. Genomics Inform. 2014. PMID: 25317108. The impact of transposable elements on gene functionality and genomic structural variations. Genesis. 2020. PMID: 33230956. The impact of transposable elements on mammalian development. Development. 2016. PMID: 27875251. The important contribution of transposable elements to phenotypic variation and evolution. Curr Opin Plant Biol. 2022. PMID: 34883307. The Intricate Evolutionary Balance between Transposable Elements and Their Host: Who Will Kick at Goal and Convert the Next Try? Biology (Basel). 2022. PMID: 35625438. The Mobilome of Reptiles: Evolution, Structure, and Function. Cytogenet Genome Res. 2019. PMID: 30739120. The power of “controllers”: Transposon-mediated duplicated genes evolve towards neofunctionalization. J Genet Genomics. 2023. PMID: 37068629. The role of transposable elements in aging and cancer. Biogerontology. 2023. PMID: 37017895. The role of transposable elements in health and diseases of the central nervous system. J Neurosci. 2013. PMID: 24198348. The Role of Transposable Elements of the Human Genome in Neuronal Function and Pathology. Int J Mol Sci. 2022. PMID: 35628657. The Structural, Functional and Evolutionary Impact of Transposable Elements in Eukaryotes. Genes (Basel). 2021. PMID: 34203645. Transposable Element Dynamics and Regulation during Zygotic Genome Activation in Mammalian Embryos and Embryonic Stem Cell Model Systems. Stem Cells Int. 2021. PMID: 34691189. Transposable Element Mediated Innovation in Gene Regulatory Landscapes of Cells: Re-Visiting the “Gene-Battery” Model. Bioessays. 2018. PMID: 29206283. Transposable Elements and Human Diseases: Mechanisms and Implication in the Response to Environmental Pollutants. Int J Mol Sci. 2022. PMID: 35269693. Transposable elements and their role in aging. Ageing Res Rev. 2023. PMID: 36773759. Transposable elements as a potent source of diverse cis-regulatory sequences in mammalian genomes. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32075564 Transposable Elements as a Source of Novel Repetitive DNA in the Eukaryote Genome. Cells. 2022. PMID: 36359770. Transposable elements as essential elements in the control of gene expression. Mob DNA. 2023. PMID: 37596675 Transposable elements as genetic accelerators of evolution: contribution to genome size, gene regulatory network rewiring and morphological innovation. Genes Genet Syst. 2020. PMID: 31932541. Transposable elements as new players in neurodegenerative diseases. FEBS Lett. 2021. PMID: 34626428. Transposable Elements Co-Option in Genome Evolution and Gene Regulation. Int J Mol Sci. 2023. PMID: 36768929. Transposable elements in Drosophila. Mob Genet Elements. 2017. PMID: 28580197. Transposable Elements in Pluripotent Stem Cells and Human Disease. Front Genet. 2022. PMID: 35719395. Transposable elements shape the evolution of mammalian development. Nat Rev Genet. 2021. PMID: 34354263. Transposable elements, genome evolution and transgenerational epigenetic variation. Curr Opin Genet Dev. 2018. PMID: 29525544. Transposable elements: all mobile, all different, some stress responsive, some adaptive? Curr Opin Genet Dev. 2018. PMID: 29705597. Transposable elements: genome innovation, chromosome diversity, and centromere conflict. Chromosome Res. 2018. PMID: 29332159. Transposable Elements: Major Players in Shaping Genomic and Evolutionary Patterns. Cells. 2022. PMID: 35326499.